
An Economics of Flexibility
“… mere purposive rationality unaided by such phenomena as art, religion, dream, and the like, is necessarily pathogenic and destructive of life; … its virulence springs specifically from the circumstance that life depends upon interlocking circuits of contingency, while consciousness can see only such short arcs of such circuits as human purpose may direct.” [1]
This is the fourth in a sequence of posts on the anthropologist Gregory Bateson originally posted on hyponymo.us (one, two, three).
What is the value of the natural world? The question seems unanswerable on the face of it — without fresh water, fertile soil, mineral deposits, solar energy, and adequate sinks for waste, no one would be around to do the valuing — but attempting to answer unanswerable questions is often revealing and worthwhile.
To do so you might make a list of every natural resource you can think of and add up the market value of the amount of that resource extracted over a period of time. Since the value provided by nature is not entirely captured by markets — no individual, corporation, or government pays money for the water treatment properties of coastal wetlands or indeed for the oxygen in the air we breathe — this initial estimate would undercount the true value. To compensate you might add an estimated shadow price or willingness to pay for replacement services as the quality or quantity of natural resource changes.
A 1997 study by Costanza et al attempted to do just that, estimating that global ecosystem services provide at least US$33 trillion to the economy each year (in 1994 US$, equivalent to about US$54 trillion in 2016). This is roughly equivalent to 20 Iraq Wars each year.
Most of the difficulty in estimating the true value of an ecosystem lies in its complexity, in two senses of the word. First in the sense of inner workings that are hard to see and understand, and second in the difficulty of duplicating the system. We must be careful when speaking of the replaceability of, say, a hectare of wetlands. An economist might tally up the “services” rendered by the hectare and find another way to render these services, possibly with mines, factories, agriculture, or other technologies. This is a weak notion of replaceability, one that I argue is inadequate to account for the value of the hectare.
Costanza et al 1997 anticipates this objection and others, and indeed they consider their result an underestimate. They call out 12 distinct limitations in the paper (section “Sources of error, limitations and caveats”), including valuing ecosystem functions at current use levels rather than at sustainable levels, and using a simplified model that assumes independence across ecosystem functions. Each limitation results in a conservative estimate, making the likely true value considerably higher.[2]
Nature as a whole may be priceless, yet parts of nature — acreages of wetlands or rainforest, populations of bees or birds — do have value that can be thought of as the cost of replacing the services they provide. I argue that such a perspective vastly undercounts the intrinsic value of the resource. I’m not speaking merely of some aesthetic sense of the unequalled beauty of nature or the purity of untouched wilderness. I’m speaking of a practical, tangible, valuable-to-humans sort of value.[3]
To more fully appreciate what’s involved in the genesis of complex hard-to-replace systems it’s worth taking a closer look at the relationship between the dynamics that describe how the system maintains stability and the slower processes by which these systems change over time.
Somatic Change and Evolution
You might recall from high school biology the debate between the Lamarckian theory of inheritance of acquired characteristics and Darwin’s theory of natural selection. The distant ancestors of the giraffe stretched their necks to reach their food, Lamarck would say, and the longer necks thus acquired were passed down to their offspring. Darwin says rather that pregiraffes who happened to have longer necks due to random mutation were more likely to survive and pass down their genetic material.
Natural selection won out, of course, and rightly so. However the common view of natural selection is simplistic, and as usual things are both more complex and more interesting than mere “survival of the fittest” and random variation. Bateson, writing in 1963, was unsatisfied with contemporary models of evolution, particularly their failure to integrate the interplay between fast, individual adaptation with slower genetic changes.
Bateson focuses on these fast and often reversible adaptations — his term is somatic changes to distinguish them from changes in the germ line — changes that occur within an organism’s lifetime in timescales of seconds to years. The concept is a familiar one: muscles grow larger with exercise, skin darkens with sun exposure, and with training we can learn concepts and skills. These variables are not directly encoded in our genes. Instead, our genes specify a homeostatic control system that itself determines, with influence from the environment, the final value of the variable.
Bateson argues that somatic change is necessary to weather environmental stress, mutational load, parasitic load, etc. Even beneficial genetic changes are stressors that the organism must adjust to in real time:
“The individual organism is a complex organization of interdependent parts. A mutational or other genotypic change in any one of these (however externally valuable in terms of survival) is certain to require change in many others — which changes will probably not be specified or implicit in the single mutational change of the genes. A hypothetical pregiraffe, which had the luck to carry a mutant gene “long neck,” would have to adjust to this change by complex modifications of the heart and circulatory system. These collateral adjustments would have to be achieved at the somatic level. Only those pregiraffes which are (genotypically) capable of these somatic modifications would survive.” [4]
The question then becomes, how was natural selection able to find a path from pregiraffe to modern giraffe, making all the necessary changes along the way? Particularly when the pregiraffe’s cardiovascular system is perfectly capable of accommodating small changes in neck length, just by working a little harder.
Bateson argues that as genetic changes accumulate, even adaptive ones, the flexibility of the organism’s somatic system is limited. The additional load from the long neck imposes a limit on the heart’s output; in emergency situations the pregiraffe’s cardiovascular system runs out of runway and fails. Bateson puts it this way:
“These considerations lead to a classifying of both genotypic and environmental changes in terms of the price which they exact of the flexibility of the somatic system. A lethal change in either environment or genotype is simply one which demands somatic modifications which the organism cannot achieve….
We face an economics of flexibility which, like any other economics, will become determinative for the course of evolution if and only if the organism is operating close to the limits set by this economics.” [5]
An example of this principle in action is the position of mammalian noses and the set of behaviors and reflexes they exhibit around water, particularly at the crucial moment when the young animal must learn to swim.

Timothy Gallwey observes in The Inner Game of Tennis:
“Once when I was walking through the San Diego Zoo, I had the chance to observe a mother hippopotamus giving her baby what looked to be its first swimming lesson. At the deep end of the pool one hippo was floating with just its nose above the surface. Soon it submerged and sank to the bottom, where it seemed to rest for about twenty seconds before pushing off with its hind legs and rising again to the surface. Then I watched a mother hippo, which had been nursing her baby in the sun, get up and begin to push it toward the pond with her snout. When the baby toppled in, it sank like a rock to the bottom and stayed there. Mother sauntered casually to the shallow end of the pool and waded in. About twenty seconds later she reached the baby and began to lift it upward with her nose, sending it toward the surface. There the young student gasped a breath and sank again. Once again the mother repeated the process, but this time moved off to the deeper end of the pool, somehow knowing that her role in the learning process was finished. The baby hippo inhaled on the surface and sank again to the bottom, but after some time, it pushed itself toward the surface with its own hind legs. Then the new skill was repeated again and again.” [6]
It’s easy to see the selective pressures at work: there have no doubt been baby hippos who did not survive their first swimming lesson.
The nasal drift of whale evolution takes this to an extreme. Breathing for a whale requires a minimum of effort because of the position of the blowhole, yet the change comes at a cost: blowholes are poorly adapted to exploring the environment through a sense of smell. Adaptations often exhibit this tradeoff between efficiency and flexibility.
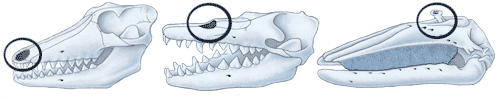
Genetic adaptations, because they are effectively set in stone, are both more efficient and less flexible than somatic adaptation. On the other hand the thing that gives somatic adaptation its flexibility, the characteristic of being reversible, makes it poorly suited to handling constant pressures — what we now refer to as “chronic stress.” In the face of such pressures, we might expect to see genetic mutations taking over for what was once homeostatically controlled:
“If the hypothetical pregiraffes carrying the mutant gene “long neck” could also get the gene “big heart,” their hearts would be enlarged without the necessity of using the homeostatic pathways of the body to achieve and maintain this enlargement. Such a mutation will have survival value not because it enables the pregiraffe to supply its elevated head with sufficient blood, since this was already achieved by somatic change — but because it increases the overall flexibility of the organism, enabling it to survive other demands which may be placed upon it either by environmental or genotypic change.” [7]
Because a certain degree of flexibility is adaptive, nay, necessary for survival, this kind of substitution should predictably occur following costly “long neck”-like mutations. In other words, Lamarckian inheritance does not happen directly, but instead is simulated by the interplay between somatic changes, genetic changes, and environmental pressures. An early form of this idea was proposed by J. M. Baldwin in 1896 and is now known as the Baldwin effect (perhaps not coincidentally Baldwin was a contemporary of Gregory Bateson’s father William Bateson, also a geneticist).
It’s worth noting that the flexibility afforded by somatic change makes for a much easier problem for natural selection to solve, since the optimization space is now amenable to hill climbing. This flexibility provides some wiggle room between the “long neck”-like stressor and the problem of cardiovascular capacity, and as a result each variable can be optimized almost independently from the other. Another way of looking at it is by imagining a cart traversing a diagonal path along a zigzagging trajectory: the wider the path the less likely the cart rolls off the edge.
Flexibility Budgets
The economics of flexibility and somatic adaptation, Bateson argues, is multiplicative. What this means is that with each additional stress, the organism’s flexibility (roughly, the range of possible environments it can survive) is reduced by some factor, say 30% or 50%, and is quickly overwhelmed as stressors accumulate.[8] This is in contrast to the economics of a household budget: because money is fungible — it can be as easily spent on a vacation as on a month’s groceries — budget stresses are merely additive.
A given adaptation or stressor will have an additive component and a multiplicative one. Additive costs are the result of drawing down a fungible resource like calories (or money), while multiplicative costs result from overlapping homeostatic signalling pathways influencing multiple variables — like contention on a conference call or on an ethernet bus, the activation of one pathway limits the simultaneous activation of the others.
Another way for systems to shift in the direction of additive (thus, more manageable) costs is to introduce redundancy and modularity. This is a particularly attractive approach for designed systems, because in addition to being made more flexible, modular systems are also easier to understand.[9] Modularity in design is discussed in more detail in this post.
The flexibility afforded by redundant biological systems — when multiple signalling pathways independently control the same variable — is the reason for the “defense in depth” described in the previous post.
Similar flexibility concerns exist at the population level. Genetic diversity is often quite high in wild populations, despite superficial phenotypic similarity. This diversity represents a storehouse of possible responses to environmental challenges, a kind of latent flexibility of the population as a whole. Crops grown in monoculture lose this flexibility, again for the sake of efficiency in optimizing certain desirable traits. As a result they are more susceptible to disease and are unable to support the kinds of complex ecosystems that regularly occur in nature.
The tradeoff between flexibility and efficiency shows up in all kinds of systems, from the corporate organizations of John Gall’s tongue-in-cheek Systemantics to the thriving neighborhoods of Jane Jacobs’s grand oeuvre The Death and Life of Great American Cities, themselves complex ecosystems that cannot be easily engineered into new urban planning projects, the litany of failed attempts through the 20th century attesting to this difficulty. Perhaps the most direct analogue of the biological Baldwin effect is the modern state, with constitutions and common law playing the role of genotype, while institutions, statutes, and elections embody somatic change.
Preimages of One-Way Functions
Up to this point it might be hard to see what calculating shadow prices of ecosystem services has to do with evolution. But there is a core idea that unites these examples, one having to do with an intrinsic aspect of value. One that, were I of a more gnomic temperament, I would vaguely gesture at, leaving the work of understanding to the few readers willing to make the effort. And perhaps if you’re one of those readers inclined to figure things out for yourself, you might pause here and guess what I’m driving at.
I am not of the gnomic school. Indeed, excessively vague pronouncements make me uncomfortable; I think rather that trying to understand difficult ideas by making them explicit is almost always better than throwing up one’s hands at their seeming ineffability. So I’ll try to articulate the core idea.
In a nutshell, this ubiquitous source of value consists of what you might think of as solutions to puzzles. More technically, given a success function that maps states in a high-dimensional configuration space to a binary success or failure (or to degrees of success), value lies in states that map to success. This function is typically one-way — difficult to invert — which means that finding these successful states is inherently costly. The function may be purposely constructed, or it may be an implicit feature of a self-assembling complex system.

For biological organisms, the success function takes genotype plus environment and produces a viability score. Most possible genotypes fail; a few succeed. Natural selection finds a subset of these few. Populations with high genetic variation can produce large numbers of successful genotypes.
Similarly, most possible configurations of natural materials do not make a stable ecosystem. Even knowing what we want out of a piece of land requires inverting the success function, most likely by cribbing from what we know works elsewhere. City neighborhoods have a success function — livability — and are only viable when certain poorly understood dynamics hold, as does architecture and design. Modern stores of value like Bitcoin are the epitome of this principle, an instance of manufactured scarcity through the privileging of semi-arbitrary mathematical properties. The underlying Hashcash algorithm literally specifies a one-way function and a success criterion.
Such Short Arcs
We humans have a sketchy track record building and maintaining systems that reliably support the general welfare. Most such efforts fail; nearly every one falls short of its promise. We have had some success, though, and thanks to Bateson and others we have the beginnings of a framework for understanding what makes these successful efforts function. The best of these systems embody what Bateson calls “wisdom,” a reliance on a dynamic whole that no single actor can fully control or even see its extent.
This framework might point us in the direction of our long term survival. Large-scale agribusiness may represent the epitome of efficiency, but home agriculture (as with WWII victory gardens) represent a flexibility that has an economy of its own. (For another perspective on designing systems while navigating flexibility–efficiency tradeoffs, see Venkatesh Rao on Fat Thinking and Economies of Variety.)
Understanding and building out this framework is becoming increasingly important. Until now we’ve had the luxury of letting systems evolve, the defunct and dysfunctional failing and fading, replaced by the continuing success of the successful. In a globalized, hyperconnected world the drive towards efficiency has produced systems that are too big to fail. If we are to continue to give birth to these global systems, it is now imperative that we learn how to get them right the first time. This will be the theme of the final post in this sequence.
Ultimately we must face the limits of our understanding as individuals. We must accept that our best-intentioned efforts may have consequences that run counter to our values. We must act with incomplete information or not at all. We must learn faster and more thoroughly; we must find ways of working better and seeing farther; we must build communities and organizations that stably and legibly represent our values.
Perhaps we will find that in time, with skillful stewardship and application of wisdom, natural resources do not degrade but appreciate.
[1] Gregory Bateson, Steps to an Ecology of Mind, “Style, Grace, and Information in Primitive Art,” p. 146
[2] Another difficulty is that it is not so straightforward to extrapolate the total value of nature from the amount extracted in a year, particularly when the renewability of these resources varies with speed of extraction and the type of resource. Economists have been tackling simplified forms of this question at least since Hotelling 1931, but even these efforts leave something to be desired, either employing too many simplifying assumptions or blurring the line between predicting commodity prices and assessing human welfare, or both. Further, efficiencies of use can change, as with the invention of improved fuel-efficiency internal combustion engines, or the particulars of use might change, as precious metals might be used for jewelry or for electronics or for hitherto unimagined purposes.
[3] For another perspective on pricing nature, found during the writing of this piece, see http://nautil.us/issue/15/Turbulence/how-to-price-a-forest-and-other-economics-problems.
[4] Gregory Bateson, Steps to an Ecology of Mind, “The Role of Somatic Change in Evolution,” p. 348
[5] Gregory Bateson, Steps to an Ecology of Mind, “The Role of Somatic Change in Evolution,” p. 349
[6] Timothy Gallwey, The Inner Game of Tennis
[7] Gregory Bateson, Steps to an Ecology of Mind, “The Role of Somatic Change in Evolution,” p. 353
[8] Consider a naive model: even if an organism has 10 times as much flexibility as it needs to reliably survive, only four mutations or stressors each costing 50% will be sufficient to kill the animal.
[9] Modular systems are never perfectly so, and are always susceptible to some degree to the problems of leaky abstractions and moral hazard.
Originally published at hyponymo.us on October 8, 2016.

